In classic wuxia fiction, poison masters are among the most feared figures in the martial arts world, wielding scorpions, venomous snakes, centipedes, and spiders as lethal weapons. Nature has evolved its own masters of venom—including scorpions, cone snails, and bullet ants (Figure 1). While their venoms function as powerful weapons for predation and defense, they also represent highly sophisticated molecular arsenals shaped by hundreds of millions of years of evolution.

Figure 1. Nature's masters of venom: scorpions, cone snails, and bullet ants (from left to right)
The neurotoxins in these venoms act as highly specialized “molecular hackers,” precisely targeting voltage-gated ion channels (VGICs), the fundamental electrical switches of excitable cells, thereby rewiring neuronal signaling.
How do these natural "molecular hackers" manipulate nervous system function? What distinct strategies do different toxins employ to regulate neuronal activity? Addressing these questions will not only advance our understanding of the fundamental principles governing nervous system function but also provide important insights for developing therapeutics for neurological disorders.
On June 10, 2026, a team led by Nieng Yan, Xiao Fan, and Jian Huang at the Shenzhen Medical Academy of Research and Translation (SMART) published a research article in Nature titled "Diverse binding poses of agonistic neurotoxins on human Nav1.6". Focusing on the voltage-gated sodium channel Nav1.6, a key regulator of the human nervous system, the researchers determined the structures of Nav1.6 in complex with three agonistic peptide toxins: the scorpion toxin Cn2, the cone snail toxin ι-RXIA, and the bullet ant toxin Pc1a (Figure 2). The study systematically reveals the previously unrecognized binding modes and mechanistic diversity among these modulators.

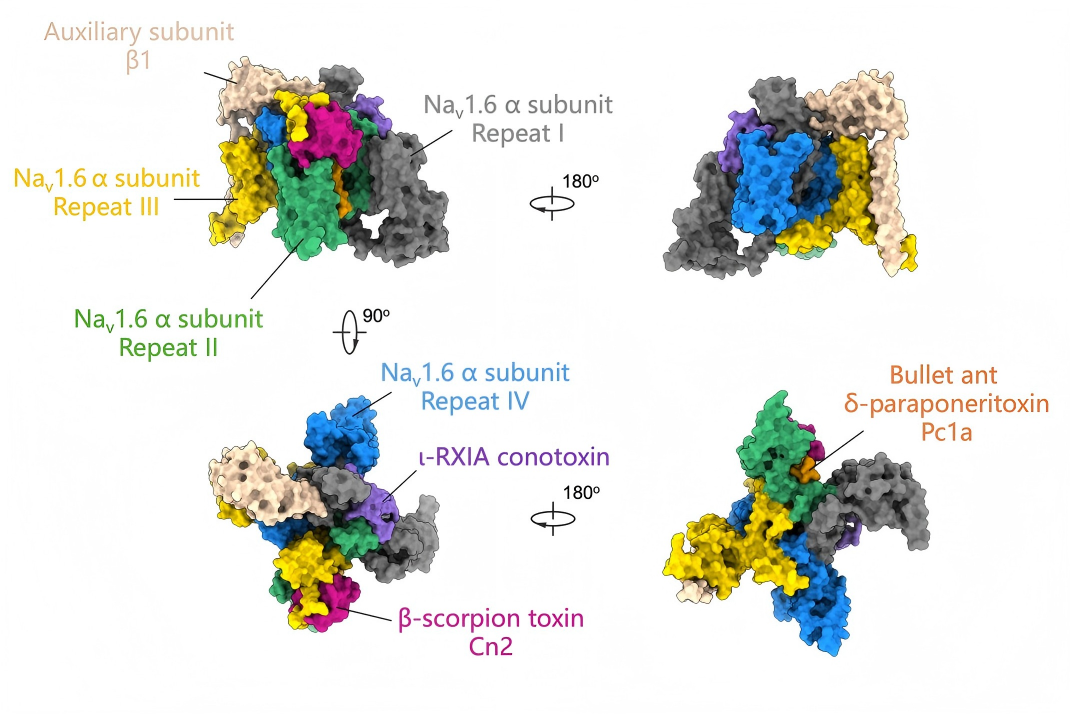
Figure 2. Diverse binding modes of Nav1.6–toxin complexes
Action potential initiation and propagation in neurons depend on the precise gating of voltage-gated sodium (Nav) channels. These channels function as electrically controlled gates embedded in the cell membrane. Upon depolarization, the voltage-sensing domains (VSDs) detect changes in membrane voltage and undergo conformational rearrangements that trigger opening of the central pore domain (PD). Sodium ions then rapidly flow into the cell along their electrochemical gradient, initiating an action potential and generating neural signals that propagate throughout the nervous system.
Among the nine Nav subtypes in humans, Nav1.6 plays a particularly prominent role in excitatory signaling. It is widely expressed in both the central and peripheral nervous systems and is highly enriched at the axon initial segment (AIS) and nodes of Ranvier—critical sites for action potential initiation and saltatory conduction. Dysfunction of Nav1.6 is closely associated with a variety of neurological disorders, including developmental and epileptic encephalopathy (DEE), making it an important target for precision therapies. Recent efforts in precision medicine have produced Nav1.6-targeting drug candidates, such as Relutrigine (PRAX-562) developed by Praxis Precision Medicines, which selectively suppresses pathological hyperexcitability associated with refractory epilepsy.
Achieving more precise and safer therapeutic modulation of Nav1.6, however, requires a deeper understanding of its natural regulatory mechanisms. As highly evolved modulators of ion channels, neurotoxins provide a unique window into the functional regulation of Nav1.6.
While prior studies have largely focused on inhibitory toxins that suppress Nav channel activity, agonistic toxins that stabilize open or activated states remain less well understood. In this study, three structurally and functionally distinct agonistic toxins were investigated: Cn2, ι-RXIA, and Pc1a. Although all target Nav1.6, they engage the channel at distinct sites and employ fundamentally different allosteric mechanisms.
Cn2: A Scorpion Toxin Anchored by an Extracellular Glycan
Nav1.6 consists of a central pore surrounded by four voltage-sensing domains. The scorpion toxin Cn2 binds between the second voltage-sensing domain (VSDII) and the extracellular loop of pore domain III.
Notably, its binding is not mediated solely by protein–protein interactions. Instead, a channel-associated N-linked glycan forms a critical component of the binding interface. This glycan is a naturally occurring post-translational modification of the channel protein. In the apo state, this glycan is flexible and poorly resolved. Upon Cn2 binding, however, it becomes stabilized through simultaneous interactions with both the toxin and the channel, allowing multiple sugar residues to be clearly resolved in the cryo-EM density map. These observations demonstrate that the glycan is not merely a passive decoration but an active participant in toxin recognition (Figure 3).
The results further show that glycans and glycan-mediated allosteric effects create an additional binding surface on the channel. This finding suggests that surface glycosylation may function analogously to a molecular glue during ligand recognition and highlights glycosylation sites as promising targets for future drug design.
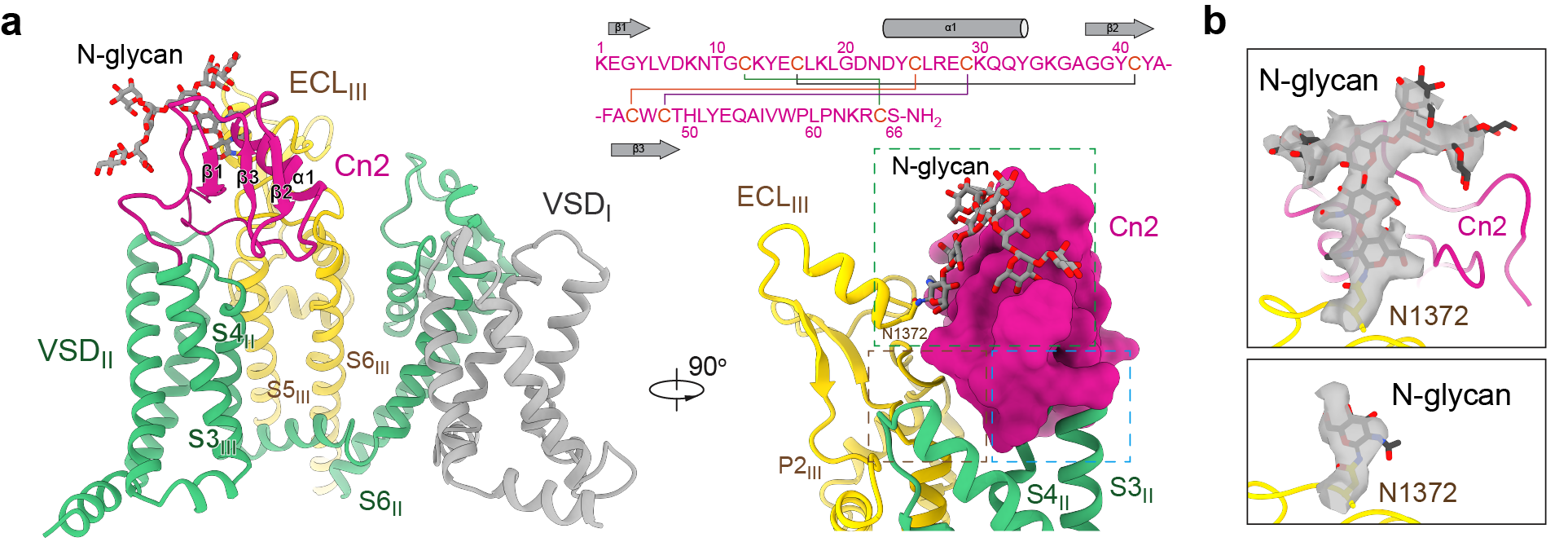
Figure 3. An N-glycan serves as a co-receptor for Cn2 binding
ι-RXIA: A "Molecular Rope" Spanning the Channel Surface
The second toxin, ι-RXIA, is derived from cone snails. Unlike the compact, nearly globular Cn2, ι-RXIA adopts an extended conformation, stretching across the channel surface like a molecular rope to tether the first voltage-sensing domain (VSDI) to the fourth voltage-sensing domain (VSDIV) (Figure 4).
The C-terminal carboxyl group of ι-RXIA engages charged residues in VSDI, stabilizing the activated state of the channel and facilitating opening. More importantly, the structural analysis, together with molecular dynamics simulations, resolved a longstanding question in the field: why does the rare D-phenylalanine residue at the third position from the C-terminus of ι-RXIA exhibit substantially higher activity than its naturally occurring L-isomer?
The answer lies in its unique chirality. The D-configuration enables the toxin to adopt a more favorable conformation, allowing its C-terminal carboxyl group to engage Nav1.6 with a more optimal geometry and thereby enhancing toxin binding and activity.
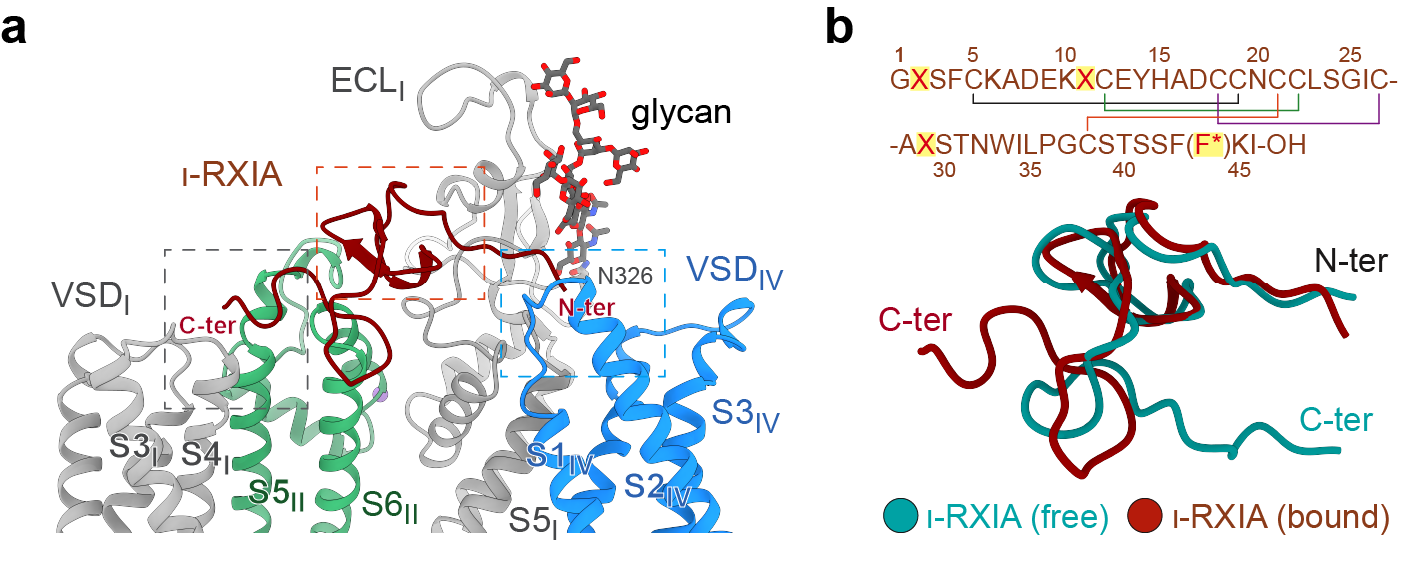
Figure 4. "Molecular Rope": ι-RXIA tethers two voltage-sensing domains
Pc1a: A Membrane-Embedded "Molecular Wedge"
The third toxin, Pc1a, is the principal neurotoxin found in the venom of the notorious bullet ant, one of nature's most painful stinging insects. Unlike Cn2 and ι-RXIA, Pc1a does not bind on the exposed surface of the channel. Instead, it inserts directly into the lipid interface of the cell membrane, acting as a molecular wedge that becomes lodged between the second voltage-sensing domain (VSDII) and pore domain III (Figure 5).
This unique binding position enables Pc1a to simultaneously contact several key structural elements involved in electromechanical coupling, including S4II, the S4–S5II linker, S5III, and S6III. Functioning much like a molecular glue, Pc1a holds the voltage-sensing domain and the pore domain together, thereby altering the conformational transitions that govern channel opening and closing.
Functionally, Pc1a not only markedly lowers the voltage threshold required for channel activation, making Nav1.6 easier to open, but also almost completely abolishes the rapid inactivation of the sodium channel, allowing persistent sodium currents to flow. This mechanism explains why bullet ant stings trigger sustained high-frequency firing of neurons, resulting in the intense and long-lasting pain characteristic of envenomation.
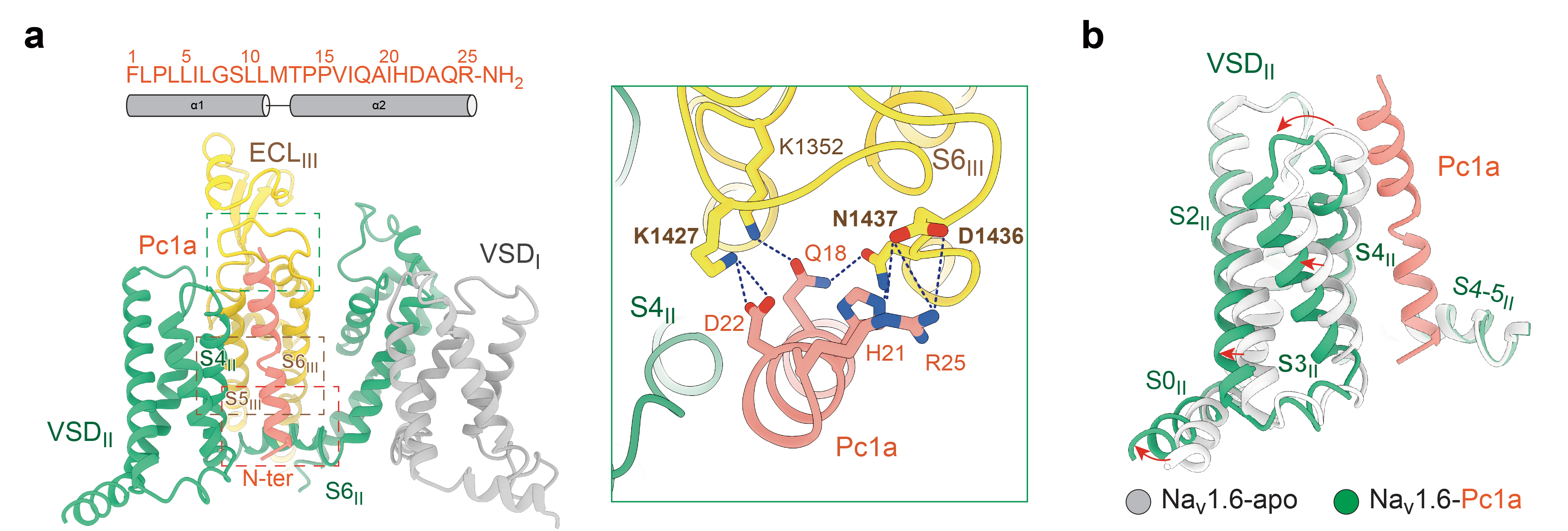
Figure 5. Pc1a restricts channel conformational changes by "gluing" together the VSD–PD
Despite their distinct binding modes, all three toxins exhibit remarkable subtype selectivity toward Nav1.6. Integrating structural data with mutagenesis analyses, the study identifies key determinants of this specificity, offering a framework for designing highly selective Nav channel modulators.
From scorpions and cone snails to bullet ants, the small molecules found in animal venoms are helping researchers decipher the fundamental language of electrical signaling in the nervous system. The three toxins employ entirely distinct modes of interaction with Nav1.6, underscoring another important lesson from nature: living organisms have long been master molecular engineers. Rather than serving solely as evolutionary weapons, these toxins may also become valuable therapeutic agents for human disease. Researchers are now gradually learning how to read the "instruction manuals" encoded within these natural molecular tools.
Nieng Yan, Founding President of Shenzhen Medical Academy of Research and Translation (SMART) and Director of Shenzhen Bay Laboratory, along with Jian Huang, Junior PI at SMART, and Xueqin Jin, Assistant Researcher at the School of Life Sciences, Tsinghua University, are the co‑corresponding authors of this paper. Xiao Fan and Jian Huang, Junior PIs at SMART, and Lin Yang, Postdoctoral Fellow at the School of Life Sciences, Tsinghua University, are the co‑first authors. Postdoctoral Fellows Jiaofeng Chen, Huan Wang, and Jinli Geng; Ph.D. student Qinglin Wu; Research Assistant Qinmeng Guo from Tsinghua University; as well as Xiaoshuang Huang, Associate Researcher at SMART, Postdoctoral Fellow Zilin Shen, Ph.D. student Fangzhou Lu, and Research Assistant Yuzhen Xie from SMART, provided significant assistance to this study. The Structural Biology Core Facility and High‑Performance Computing Platform at SMART provided key technical support. This work was supported by the National Natural Science Foundation of China, Guangdong Provincial Talent Programs, the Beijing Natural Science Foundation, the Beijing Advanced Innovation Center for Structural Biology, a Long‑Term Fellowship from the International Human Frontier Science Program Organization (HFSP), and the PI Start‑up Funding from SMART. The 3D models of the Nav1.6–toxin complexes were produced by the R&D and Maintenance Center of SMART. The team also expressed special thanks for support provided through the "Mindray Professorship" funded by the Pengrui Foundation.
An earlier version of this study was released on May 5, 2026, through the LTS Preprint Server (LTSpreprints.org)[1], enabling early dissemination and scientific discussion prior to peer-reviewed publication. The platform provides rapid access, broad visibility, and long-term archiving for emerging research outputs. Articles posted on LTSpreprints are increasingly being indexed by major academic platforms such as Google Scholar and ResearchGate, reflecting their growing academic impact. The platform is dedicated to fostering an open, efficient, and standardized environment for scientific communication by providing rapid dissemination, broad visibility, and long-term archiving for research outputs.
Reference:
[1] FAN, X., Huang, J., Yang, L., Chen, J., Wang, H., Huang, X., Geng, J., Wu, Q., Xie, Y., Lu, F., Guo, Q., Shen, Z., Jin, X., & Yan, N. (2026). Diverse binding poses of agonistic neurotoxins on human Nav1.6. LangTaoSha Preprint Server. https://doi.org/10.65215/LTSpreprints.2026.05.05.000226
Translation: Jianming JIA
Proofreading: Xiaoshuang Huang, Yuzhen Xie
talent@smart.org.cn
researcher@smart.org.cn
recruitment@smart.org.cn
graduate_office@smart.org.cn
graduate_admission@smart.org.cn
otl@smart.org.cn
smartfund@smart.org.cn
pr@smart.org.cn
Subscription successful! Thank you for following SMART.
